尽管生物具有惊人的多样性,但是生命的形式和过程受到普遍原理的限制,普遍原理在大小尺度上发挥作用。

老扬·勃鲁盖尔于1613年创作的油画《动物进入诺亚方舟》
宇宙中是否存在另外一种进化实验?这样的进化实验看起来会是怎样的?在查尔斯·达尔文时代,或许这些问题都是猜测,但是达尔文从未考虑过这些问题。一旦生命出现在地球上,会在全球范围内迅速繁殖,呈现出各种引人瞩目的形式,造成了非同寻常的变化,这些形式和变化跟生物圈和岩石圈有着密不可分的联系。我们呼吸的氧来源于光合作用的产物,而氧如此普遍、如此丰富,最终足以推动复杂多细胞生物圈的形成。
理解生命现象的核心是:解释清楚物理定律在多大程度上能够缩小生物结构中的所有等级层次的可能性范围。最终,要想真正地迎接这一挑战,我们可能就必须找到其他的进化实验,然后进行有效的统计对比。天体生物学激发了科学家对生命的可能替代结构进行了大量的研究,而天体生物学是围绕以下这些方面进行研究的一门科学:生命的起源、生命的进化、生命如何覆盖了地球表面,其他星球是否存在生命,以何种形式存在,等等。即使在没有观察到外星生命的情况下,我们仍然能够通过地球上的进化实验了解很多情况。
翅膀和轮子
例如,看一看老扬·勃鲁盖尔(Jan Brueghel the Elder)在其油画左边描绘的那群生物,休闲的欣赏者很容易看出,生命是无限的,生命的形式和形状只是受到想象力的限制。然而,不管观察多少遍,生命一定符合物理定律。然而,目前的科学还不知道:在物理定律约束的范围内,有多少可能的解决方案可以构建一个自我复制的体系;科学也不清楚:物理学在多大程度上限制着进化过程的产物。
在生物体系的尺度上,物理定律肯定限制了解决生活难题的工程方案。例如,考虑一下运动问题。在当今一篇典型的论文中,迈克尔·拉巴贝拉(Michael LaBarbera)论述了一个自古以来生物学家在咖啡桌旁最爱讨论的问题:为什么生物不使用轮子呢?人类在各种形式的运动中都使用轮子,为什么其他生物拒绝使用轮子?进化出能够旋转的肌肉和静脉是个生物力学难题,除此之外,轮子还有一个固有的物理问题,它受到所跨越地形的限制:如果障碍物高于轮子的半径,那么轮子就无法翻越,除非把轮子举起来。
跟体型较大的生物相比,地形对于小蚂蚁来说更加不规则。即使对于像人类这么大的动物来说,腿在翻越障碍物方面也比轮子要有效得多。此外,许多基底(如沙地或湿土)都会对轮子的运动造成相当大的阻力。有趣的是,在世界上具有干旱特征的平原地区,粪甲虫推动着几乎呈圆球状的粪球(见图1),像风滚草一样在平原地形上滚过。在快速运输能够成功的地形上,生物学的确在探索球状结构和轮式结构的应用。

图1轮子与生命。根据目前的了解,任何生命形式都没有用轮子代替腿,但在世界上地面干燥和平坦的地区,生物在进化过程中确实在探索将球状结构和轮式结构的装置作为有效运输的手段。在这里,一只粪甲虫正在把自己的财富滚到家中
人们可能会想到:在外星世界,星球的地形不可避免地存在地面缝隙、岩石断裂和凹凸不平等情况,这样就会导致生物选择的是腿而不是轮子。这种选择受制于物理规律,并不是偶然、暂时的进化巧合。
另一种运动为不可逾越的物理障碍提供了证据。有关昆虫飞行的空气动力学是生物物理领域非常成熟的学科。生物进化已经探索了大气中空气动力学升力的简单定律。翅膀上方产生低压区所需要的典型机翼状结构已经磨炼出来(见图2)。然而,这种进化的雕琢还不足以使许多昆虫飞起来,这些昆虫的笨重身体需要额外的帮助。在过去的20年中,研究显示了昆虫如何设法通过微小的翅膀逐步获取上升和推进的能力。一种这样机制叫作“后合前张(clap and fling)”:当推动翅膀向后摆动时,翅膀就拍合在一起,这一策略将翅膀间的空气排挤出去,提供了额外的推力;然后,当翅膀开始向前摆动时,翅膀张开,空气冲进来填满间隙,增加了翅膀表面的空气循环,从而提高了上升的动力。

图2昆虫翅膀。蜻蜓翅膀的结构不是偶然的进化巧合,相反,这是在物理定律引导下的进化产物。这些物理定律提供某些方案来获得可能的升力和推力
关于生命的分子限制
从飞行的鸟类到步行的蚂蚁,所有的生物都是由分子构成的……这些分子也是有限的。20世纪70年代,生物化学家开始探索构成生命的各种蛋白质。似乎跟动物学领域的同事一样,生物化学家也需要面临令人讨厌的各种可能性。20种氨基酸串在一起,形成的链长仅有200的时候,就有可能形成大约10260个蛋白质;增加这个链的长度,蛋白质的数量会更加令人难以置信。为了理解所有这些分子,需要多少世纪的生物化学研究工作呢?然而,随着研究人员对蛋白质进行测序和研究其折叠的方式,这一点变得明晰起来:不管氨基酸的序列是怎样的,蛋白质分段呈现出的形状数量的确是非常有限的(见图3)。
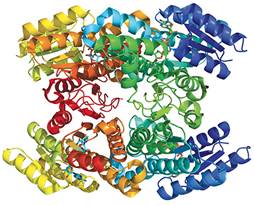
图3有限的蛋白质结构。丁二醇脱氢酶参与跟特定有机化合物的氧化还原反应。尽管蛋白质具有复杂性,但它主要是由柔性氨基酸串连接的α-螺旋结构(弯曲的带状结构)和β-折叠结构(扁平的箭头结构)组成的。其结构说明了蛋白质是如何由简单的基序组合而成
把蛋白质分解成单独的部分,就会呈现出四种主要的折叠方式。被称为α-螺旋的结构是氨基酸的右螺旋折叠方式,由一个氢键连接到一起,该氢键位于一个氨基酸的氨基氢和羧基(-COOH)之间,而该氨基酸位于序列中三四个氨基酸之前。被称为β-折叠的折叠结构是通过氢键连接起来的平行氨基酸长链。第三种和第四种折叠方式是混合基序,是由氨基酸上分别呈现的螺旋结构和折叠结构组合而成的。一般的螺旋结构和折叠结构的混合结构被称为α+β,交替出现的螺旋结构和折叠结构被称为α/β,后者为特殊情况。被称为TIM(磷酸丙糖异构酶)桶状基序、夹层状基序和卷状基序的子类界定了螺旋和折叠组合的特定方式。
对于折叠组合方式的有限性,一个解释是:在生存的斗争中,特定的折叠形式足以形成有竞争力的生命个体,这些形式已经在早期的进化中锁定于生物体内。拿建造房子作类比可能有助于澄清上述观点:你不用到建筑商的院子,也无须利用所找到的各种砖,只需挑选出能够建造房子的几种砖即可。一旦你确定这些砖能够建造一所好房子,你就会批量生产这样的砖。
虽然偶然性论点可能具有说服力,但基本的物理定律有可能在选择蛋白质特定形式的过程中起到了作用。跟分子一样,氨基酸链往往用这样的一种方式进行折叠,以达到最低的能耗状态。每个连续的折叠步骤由热力学原理驱动,以达到最稳定的状态。这些折叠步骤不是独立的,折叠蛋白质的不同部分会对该蛋白质其他部分的折叠模式产生影响。蛋白质会寻求有限数量的热力学上有利的位形。
长期以来,一些生物学家一直在争论:生物学定律是否存在?从达尔文进化论的角度看待生物领域是否更好?在达尔文的进化论中,秩序不是预先注定的,变异和选择决定了生物可能性的广阔前景。这两个观点不仅是相容的,而且是不可分开的。达尔文的进化论通过变异和选择测试了多样的生命形式,但产生的生物符合物理定律,严格受到普遍原理的约束。例如,达尔文的进化论产生了大量的蛋白质,这些蛋白质对于选择的各种结构和功能是有用的。然而,热力学大大限制了构成分子的基序数量。
生命元素
物理学原理可能会限制构成生命的每个元素。富有想象力的科学家想到了用来替代生命的各种化学物质,其中一些普遍的化学物质基于硅元素。硅的姊妹元素――碳,在元素周期表中正好位于硅的上方,和碳相比,硅有一套额外的电子轨道。因此,在某种程度上来说,硅比碳更活跃,形成稳定长链结构的能力较差,不像数以百万计的碳化合物那样。一个例外情况是,硅形成硅酸盐的能力很好,硅酸盐是跟氧结合而形成的极其稳定的结构。
自从45.4亿年前我们的地球形成以来,硅化学实验一直在地球上自然地进行着,生成的硅基化合物的多样性令人印象深刻。但是,这些硅基化合物都是矿物、玻璃和各种非晶态结构,它们共同构成岩石。图4显示了一些硅基化合物。实验室的研究已经产出了复杂的硅化合物,这些硅化合物不太像岩石,而是更像构成生命的材料。例如,倍半硅氧烷是一类有机硅化合物,形成了链状和笼状结构,给人印象深刻,使人产生了硅碳混合生命的想法。然而,据我们所知,有机硅化合物的复杂性和多样性非常有限。碳原子的大小(使其能够形成稳定的单键、双键和三键)和能力(在极少能量损耗和释放的情况下,能够跟许多元素结合形成键,在一系列元素之间进行键切换)使碳元素有利于多种分子的结合。
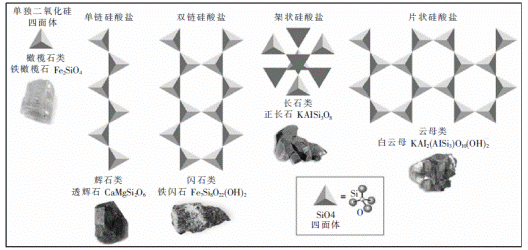
图4硅结构。跟氧结合,硅可以形成多种的硅酸盐结构。硅酸盐是二氧化硅四面体的不同组合,而二氧化硅四面体是通过普通的氧原子连接的。硅酸盐极为规则,通常呈结晶状,更适合形成岩石,不适合形成生命。这里显示的是硅酸盐的一般结构,每个结构类别中都有特定类别实例,特定的矿物是每个类别的代表。小插图显示了四面体的示意图,中心为硅原子,顶点处朝向外面的是氧原子。在硅酸盐原理框架图中(右四),填充的三角形代表四面体,凸起的氧原子是朝向页面里面的
上述情况决不局限于地球上。天体化学研究的是宇宙的化学性质,跟天体生物学有交叉。在过去的20年中,天体化学领域的发现引人瞩目。
在银河系各处,包括四处弥漫的星际云、巨型分子云和形成新恒星系统的原行星盘,碳基化合物正在合成。其中有些化合物是简单的,如NH2CN、CH3SH和CP等分子种类,但是也会产生许多更加复杂的化合物。多环芳烃像一张邮票一样,具有重复单元的复杂连接结构,在这种情况下重复单元就是芳环。研究人员认为:多环芳烃最终能够形成管状结构、巴克球结构和类似洋葱的层状结构。更重要的是,研究人员发现了异丙基氰和其他的含氰化合物。氰化物类可能对有益于生物形成的分子起着相当于前体的作用,这类分子有着分支结构,类似于对生物很重要的分子,如氨基酸。由于空间的物质密度低和寒冷,研究人员认为:这些化合物会形成于硅质颗粒和碳质颗粒上,这些颗粒的表面能够将反应物聚在一起。在某些情况下,星际辐射对反应能够起到催化作用。
最有趣的是,研究人员从降落在地球上的陨石中获得了碳化合物。一类陨石为碳质陨石,其中包含我们太阳系一些最原始的物质材料。研究人员发现:碳质陨石中包含构建碳水化合物的糖、构成蛋白质的氨基酸、浸泡于水中时性质类似于细胞膜脂的长链羧酸,甚至还包含作为遗传密码信息单位的核碱基。因此,这些证据表明,太阳系的原行星盘(诞生了太阳和行星的薄饼状尘埃和岩石漩涡)有合成生命基本单元的理想条件,这些基本单元相结合形成了生命分子的主要类别。
陨石不能提供多种多样的硅化合物。研究人员观察到过一些碳化硅,但是陨石中的硅基化学物质再次局限于矿物和玻璃。这些从太空来到地球上的信使可以证明,硅基生命和碳基生命是具有相对优势的。当然,这些陨石使得如下观点更加可信:碳基生命是普遍存在的,它并不是地球化学领域的一朵奇葩。
使碳适合作为生命原子的所有属性都基于一个简单的概念――泡利不相容原理,该原理确保加入轨道和亚轨道的电子都整齐地配对。特别是,泡利原理明确了电子的顺序层,这些顺序层决定了电子的半径和反应。不管是在温血生物中还是在原行星盘混乱的旋转气体中,碳都倾向于形成一系列氨基酸,这表明:泡利原理在基础的物理层面上支持着生物的组合,适用于任何地方。
达尔文进化论不能自由和随意挑选化学周期表中的元素,也不能创造出无穷的生命形式。复杂的碳化学是生物学的一个特征,因为它源自量子物理学定律。碳在分子产物方面具有创造多样性的独特能力,这种能力在分子云中显而易见,在蟑螂中同样显而易见。人们期待,同样的能力存在于全宇宙的多个行星环境中。
生命的能量获取机制
即使在亚原子的尺度上,我们仍然发现生命的简朴之美源于物理学。生物化学家彼得·米切尔(Peter Mitchell)首先提出了生物从环境中获取能量的详细机制,他因此获得了1978年诺贝尔化学奖。生命似乎不会轻易从核裂变中获得能量。一些人声称,假设的生命可能会利用电离辐射作为能量的来源,但是这种辐射通常会对许多分子造成严重损害。核聚变需要大量能源,无法成为生物获取能量的合理方式。在原子中,有机体可以获取能量的部分是相对容易接近的电子,在米切尔的化学渗透过程中,生命利用的正是电子。
电子从电子供体(电子供体是具有失去电子倾向的分子)形成的环境中聚集,在细胞膜或线粒体膜内穿行,在途中被电子受体接受。在你我体内,这层膜是线粒体膜,电子供体是有机物,电子受体是氧。你午饭吃的三明治是一种美味的吃电子的方式。
图5描绘了化学渗透机制。在静电势能的驱动下,电子在膜内穿行,存在于膜中的一系列蛋白质利用这些电子的能量将质子从线粒体之内移动到线粒体之外,结果是形成了质子梯度。在渗透的作用下,外部的质子倾向于返回线粒体内部,以中和质子梯度。外部质子不是通过随机扩散穿过薄膜而返回线粒体内,通常这个膜对质子来说是不可穿透的。相反,这些质子是通过一种复杂的分子装置返回内部的,这种装置是一种酶,叫作ATP合酶,是制造ATP(三磷酸腺苷)的酶。当质子通过ATP合酶的时候,会引起这种酶的组分旋转。
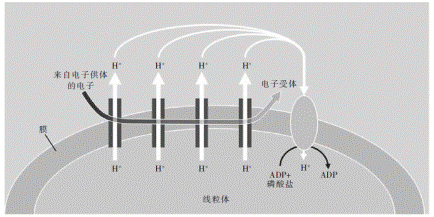
图5化学渗透过程。如正文所述,生命从有机物质或某些其他电子供体中获取电子的能量,最终将其存储在三磷酸腺苷(ATP)的磷酸键中。随着所供给的电子被运送到氧之类的电子受体中,该电子穿越部分线粒体膜(或在一些生物体中为细胞膜)。在膜中穿越时,电子经过从该电子获取能量的一系列蛋白质,这些蛋白质利用获取的能量将质子(H+)转移到线粒体之外。在渗透的作用下,排出的质子通过ATP合酶(黄色)重新进入,ATP合酶通过二磷酸腺苷(ADP)和一个磷酸基构建ATP分子。尽管所涉及的蛋白质具有生化复杂性,但该过程的物理简单性表明,这是一个获取能量的普遍机制
化学渗透的简朴之美在于:电子供体和电子受体的变换能够使生命生长于各种地方。把电子受体从氧变为硫酸盐,就可以形成硫酸盐还原菌,这种细菌可以生活在地下深处,它们是生物圈中负责硫循环的微生物。如果把夹层中的分子换成铁、氢或氨作为电子供体,那么就会得到化学合成菌,它们生活在岩石、火山热水池和热液喷口处,靠行星地质的原材料为生,而不是依赖阳光和来自其他生物体的食物。
让我们回过头来再看一下这个过程。化学渗透始于容易获取的亚原子粒子(电子),这些亚原子粒子可以提供一些能量,这些能量用来产生另外一种亚原子粒子的梯度――质子梯度。然后利用这一梯度,通过渗透的作用,产生一种有效存储电子能量的分子,便于在任何需要的地方释放。除了质子之外,生命或许能够利用离子来创建梯度。甚至在地球上,一些生物体好像也能够利用钠离子梯度的力量,但是原理是相同的。
化学渗透非常简单。那么,化学渗透是偶然的还是普遍的呢?正如我们所看到的,裂变和聚变是更加难以利用的能源;对于能量需求较少的生命来说,原子核就像一头难以驯服的野兽。假如所涉及的分子偶然发生了变化,或许利用了不同的离子创建了梯度,情况似乎仍然有可能是这样的:任何地方的生命都会利用从电子中得到的能量,而电子也相对容易从原子中剥离,完成提供能量的任务。
物理现象与偶然事件
生命一定是由物理定律塑造的。鸟类一定会符合空气动力学原理,蛋白质折叠一定符合热力学原理,利用电子获取能量的体系一定符合亚原子粒子的各种能量状态。不太清楚的情况是:物理学在多大程度上缩小了达尔文进化论的可能性范围。长期以来,人们已经了解的基础性物理学原理是:生命是通过占有主导地位的碳基分子构建的,而不是由硅基分子构建的。然而,最近科学家提出:以前似乎具有偶然性的其他选择并非随机事件,这些事件基于针对自我复制的进化体系的概率统计、能量获取和优化安排。这样的例子包括蛋白质的结构,甚至还包括生命从数百种可能性中选来创建这些蛋白质的20种氨基酸。
物理学家的一个任务是:审查不同层次的生命中偶然性存在的可能程度,探索生命系统结构的可预测性。在这个探索过程中,天体生物学家的作用是:试图确定我们是否能够发现进化实验的另一个例子,利用这样的实验验证如下的假设:从能量获取机制到整个生物体的形式和形状,生命都被严格限制在几个类别之中。在这篇文章中以及在别处,我都曾经提出过,情况的确是这样的。尽管对于许多想象力丰富的科幻作家来说是令人失望的,但是,如果这一假设最终被人们接受,我们会了解到地球上的生命非常普通。
资料来源 Physics Today
责任编辑 岳 峰
――――――
本文作者查尔斯·科克尔(Charles Cockell)是英国爱丁堡大学天体生物学教授。