一系列简单的步骤即可解释从单细胞进化到多细胞生命这一重大转变。
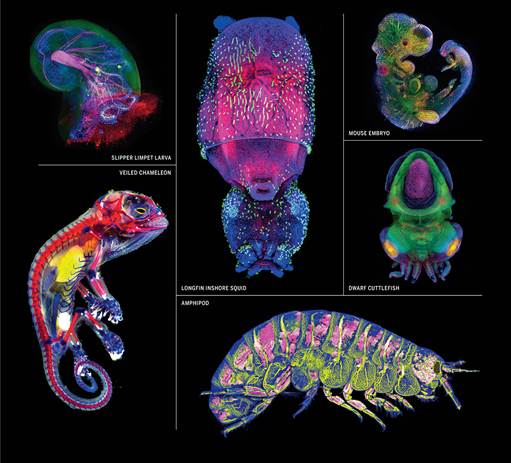
在马萨诸塞州伍兹霍尔研究所的海洋生物实验室里,学生们用不同的着色剂对动物胚胎进行了处理,多细胞的特性使得复杂结构和特殊组织能够在动物胚胎中被不同着色剂显示出来(见封二)
数十亿年前,生命跨越了一个门槛。单个的细胞开始结合起来,众多无定型的单细胞生命正逐渐演变为如今拥有各种形态和功能的多细胞生命,从蚂蚁到梨树再到人类。这是生命历史上最重要的转变,直到最近,我们仍不知道它是如何发生的。
单细胞和多细胞生命之间的鸿沟看似几乎无法逾越。单个细胞的存在是简单而有限的。就像隐士一样,微生物只需要关心自身温饱;虽然有些微生物偶尔也会团结协作,但与其他微生物之间无论协调还是合作都是不必要的。相比之下,多细胞生物体内的细胞,从某些藻类的4个细胞到人类的37万亿个细胞,放弃了自身的独立性,顽强地结合在一起;它们肩负特殊功能,为了更大的利益而缩减自身的繁殖,只增长到它们履行其功能所需的数量即止。它们一旦“造反”,癌症便会爆发。
多细胞生物带来了新的能力。例如,动物为了寻找更好的栖息地、躲避捕食者和追捕猎物而获得了移动的能力。植物可以深入土壤寻找水分和养分;它们同样也可以向阳光充足的地方生长,以期将光合作用最大化。真菌通过建立巨大的生殖结构来散播孢子。但是,赛格德匈牙利科学院生物研究中心的进化生物学家拉什路·纳吉(LászlóNagy)说,尽管多细胞生物具有诸多优势,但在传统上它被视为“一个有着巨大遗传障碍的重大转变”。
现在,纳吉和其他研究人员逐渐意识到这可能并没有那么困难。证据来自多个方面。部分生物群体的进化历史记录了从单细胞到多细胞形式的反复转变,表明这种遗传障碍并非难以逾越。简单的多细胞生物与其单细胞亲属之间的基因比对表明,细胞间结合与协调活动所需的许多分子设备可能早在多细胞生物进化之前就已经存在了。巧妙的实验已经证实,在试管中,单细胞生命可以在短短数百代内演化出多细胞生物的起源这一进化瞬间。

团藻,一种由数百或数千个细胞组成的水藻群落,为多细胞生物起源提供了线索
进化生物学家仍在争论是什么促使细胞的简单聚集变得越来越复杂,从而引出了如今生命奇妙的多样性。但是,踏上这条道路似乎不再那么令人气馁。“我们开始了解到它是如何发生的,”西雅图华盛顿大学进化生物学家本·科尔(Ben Kerr)说道:“你采取的似乎是进化的主要步骤,并将其分解成一系列小步骤。”
多细胞生命的蛛丝马迹可以追溯到30亿年前,在那时,化石记录中似乎出现了类似微生物垫的印记。一些人认为,在美国和亚洲发现的距今20亿年的、有可能是蓝绿色或绿色藻类的螺旋状化石,以及在南非记录到的被称为卷曲藻(Grypaniaspiralis)的25亿年前的微丝,是多细胞生命的第一个真实证据。其他种类的复杂生命体直到很久以后才出现在化石记录中。被许多人认为是活着的最原始生物的海绵可以追溯到7.5亿年前,但是许多研究人员认为一群5.7亿年前很常见的、被叫作埃迪卡拉的叶状生物是第一个明确的动物化石。同样,化石孢子也表明多细胞植物是在至少4.7亿年前由海藻进化而来的。
植物和动物成为多细胞生物都只经历了一次飞跃。但在其他生物种群中,这种转变却一再发生。基于对不同种类的真菌――有些是单细胞的,有些是多细胞的――相互间如何联系的回顾性研究,纳吉2017年12月8日发表在bioRxiv上的一份预印本总结道,在十几种不同的情况下,真菌很可能会以子实体的形式进化出复杂的多细胞生物――就像蘑菇一样。藻类也同样如此:红藻、褐藻和绿藻均在过去的10亿年左右进化出了它们自己的多细胞形态。
加州大学伯克利分校的生物学家妮可·金(NicoleKing)找到了一个可以展示这些古老转变的窗口:领鞭毛虫――一群似乎正处于向多细胞生物进化边缘的现存原生生物。这些动物的单细胞近亲具有鞭毛和衣领状短毛,类似于排列在海绵腔体内用来过滤食物的“衣领”细胞。有些领鞭毛虫本身可以形成球形的菌落。20多年前,金学会了培养和研究这些水生生物,到2001年,她的基因分析开始对当时的观点――向多细胞生物的转变是一个重大的基因飞跃――提出质疑。
她的实验室开始发现一个又一个基因,这些基因一度被认为是复杂动物所独有的――而且似乎在孤立的细胞中并不需要。领鞭毛虫具有编码酪氨酸激酶的基因,在复杂的动物体内,酪氨酸激酶有助于调控特殊细胞的功能,如胰腺中的胰岛素分泌。他们有细胞生长调节因子,如p53,一种因与人类癌症相关而臭名昭著的基因。他们甚至拥有编码钙粘蛋白和c型凝集素的基因,这些蛋白质可以帮助细胞粘连在一起,使组织保持其完整性。
总的来说,通过对21种领鞭毛类生物活性基因的调查,金的团队发现这些“简单”的生物体有大约350个一度被认为是多细胞动物独有的基因家族,他们将这一研究结果发表在了5月31日的eLife上。正如她和其他人所相信的那样,如果领鞭毛虫能让我们一睹动物单细胞祖先之“芳容”,那么这种生物就已经为进化成多细胞生物做好了充分准备。金和她的实验室“把原生生物放在研究动物起源的前沿。”西班牙国家研究委员会及西班牙巴塞罗那庞培法布拉大学的进化生物学家英纳基·鲁兹-特里洛(IakiRuiz-Trillo)说道。
这些基因的原始版本可能并不具备他们后来所具有的功能。例如,领鞭毛虫有对神经元至关重要的相关蛋白质的基因,但他们的细胞并不像神经细胞,金说道。同样地,它们的鞭毛中含有一种蛋白质,这种蛋白质在脊椎动物中可以帮助形成身体的左右不对称,但它在单细胞生物中的作用却是未知的。然而,无论从哪方面来讲,领鞭毛虫的基因组都不指望自己能进化成多细胞生物;它们缺乏一些关键的基因,包括像Pax和Sox这类在动物发育中至关重要的转录因子。“这些缺失的基因让我们对什么是真正的动物界革新有了更好的认识。”金说。
当细胞结合在一起时,它们并不仅仅是给现有的基因赋予新的用途。有关团藻(一种能够形成美丽的、具有鞭毛的绿色球状的水藻)研究表明,多细胞生物也找到了利用现有功能的新方法。团藻及其近亲跨越了向多细胞的过渡状态。团藻个体有500到60000个细胞排列在一个空心的球体中,而它的有些亲戚,如盘藻,仅有区区4到16个细胞;其他团藻近亲则完全是单细胞生物了。通过对比从一个细胞到数千个细胞的生物学和遗传学连续体,生物学家们正在收集生物变得越来越复杂时的需求。“这群海藻教给了我们多细胞生物进化相关的一些步骤。”亚特兰大佐治亚理工学院的进化生物学家马修·赫伦(Matthew Herron)说道。
这些研究表明,复杂生物体中特殊细胞的许多功能并不是新出现的。恰恰相反,常见于单细胞生物的特征和功能在它们的多细胞近亲中,在时间和空间上进行了重新排列,普林斯顿大学理论生物学家科瑞娜·塔尔尼塔(Corina Tarnita)如是说。例如,在团藻的单细胞亲戚衣藻中,被称为中心粒的细胞器即具有双重功能。在细胞一生的大部分时间里,中心粒锚定在两条推动细胞畅游于水中的旋转的鞭毛上。但是当细胞准备繁殖时,它会蜕去鞭毛,此时中心粒即向细胞核移动,在那里它们帮忙将分裂中的细胞的染色体拉开。之后,子代细胞分别重新长出鞭毛。衣藻既能游动又能繁殖,但二者不能同时进行。
多细胞团藻在游动的同时亦能繁殖,因为它的细胞已经分化了。较小的细胞一直具有鞭毛,鞭毛能够将营养物质扫过团藻的表面,并帮助团藻游动。较大的细胞没有鞭毛,而是专职利用中心粒进行细胞分裂。
团藻也重新定位了单细胞祖先的其他特征。在衣藻中,当夜间光合作用停止并且资源稀缺时,一种古老的应激反应通路会阻止其进行繁殖。但在团藻中,同样的通路在它的游动细胞中始终处于激活状态,以永久阻止这些细胞繁殖。科尔说,单细胞祖先对环境信号的应答被用来促进其更复杂后代的“劳工”细胞的分裂。
第三组生物体暗示了现有基因和功能的重新定位是如何发生的。在过去10年里,鲁兹-特里洛和他的同事们已经将十几种原生生物的基因组和动物的基因组进行了比对――这项比对强调了动物基因组更宏大的规模和更复杂的特性,他们将这项研究结果发表在了7月20日的eLife杂志上。但是,当鲁兹-特里洛、现供职于以色列雷霍沃特魏茨曼科学研究所的阿尔瑙·瑟比-皮德罗斯(ArnauSebé-Pedrós)以及来自巴塞罗那基因调控中心的卢西亚诺·迪克罗斯(LucianodiCroce)对原生生物Capsaspora的基因调控信号组合进行分析后,他们有了更有说服力的发现。他们发现原生生物利用与动物相同的一些分子――被称作转录因子的蛋白质分子和并不编码任何蛋白质的长链RNA分子――在特定的时间和地点开启和关闭基因。但是它的启动子――与转录因子相互作用的调节DNA――比动物的启动子要短得多,也简单得多,这就意味着不那么复杂的调控,这些研究团队2016年5月19日在《细胞》杂志中对研究结果进行了报道。
对于鲁兹-特里洛和他的团队来说,这一发现指向了多细胞生物的关键:在基因调控中增加了微调的部分。与单细胞祖先相比,这似乎是一个巨大的飞跃,但如果这在一定程度上就是个重置基因开关的问题:使现有基因在新的时间和地点被激活,这一巨大飞跃看起来也就没那么高不可攀了。佐治亚理工学院的威廉·拉特克利夫(William Ratcliff)表示:“这就是进化一直在做的事情,善用周围的事物来达到新的目的。”
这种节约型再利用也许能够解释拉特克利夫实验室中展现的迅速转变。他在实验室培养中重新创造了进化,而不是去查看化石记录或者对现有生物的基因组进行比对。他解释说:“我自己的研究并没有试图找出现实世界中发生了什么,而是研究细胞如何在进化中增加了复杂性这一过程。”
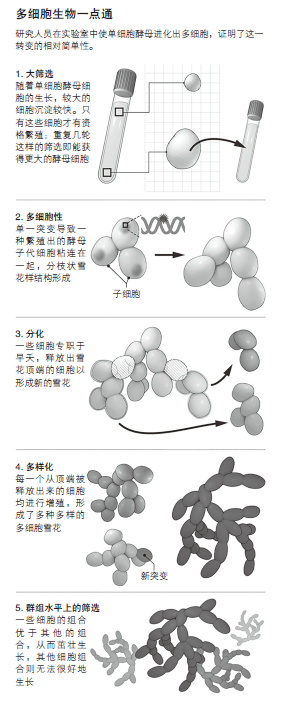
拉特克利夫是明尼苏达大学圣保罗分校的博士后,他与迈克尔·特拉维萨诺(MichaelTravisano)共事。他将酵母培养变成了人工选择的一种形式。他以细胞黏附生长在培养瓶底的速度作为衡量标准,只允许最大的细胞存活和繁殖。两个月内,随着新生的子代细胞黏附在母体上并形成分支结构,多细胞簇开始出现。(《科学》,2011年11月18日,第893页。)
随着每一代培养中的酵母的不断进化――有些已经经历了3 000多代――酵母“雪花”变得越来越大,酵母细胞也变得更加持久、更加细长,一种新的繁殖方式也随之进化。在大片的雪花状酵母中,一些沿着长的分支生长的细胞经历了某种形式的自杀,它们会释放处于顶端的细胞,开启新的雪花之旅。垂死的细胞牺牲掉自己的生命,以成全群体的繁殖。这是细胞分化的最初形式,拉特克利夫解释道。他刚刚开始探索这些快速显现的特征的遗传基础;它似乎是一种被赋予了新功能的现有基因和失效的其他基因的混合体,比如一种有助于酵母细胞分裂的基因。
酵母还衍生出了一种保护措施,这种保护措施对多细胞生物来讲至关重要:这是一种防止细胞背叛的方法。当突变使某些细胞异于其他细胞,并且可能不那么乐于团结协作时,细胞中的叛徒就出现了。在复杂生物体例如人类中,这种保护机制在某种程度上来自于拥有可摧毁异常细胞的免疫系统。它还依赖于代际间的瓶颈期,在瓶颈期时,单个细胞(例如,受精卵)可作为下一代的起点。其结果就是,新一代的所有细胞生成时在基因上都是一模一样的。雪花状酵母用自己的方式来清除自身那些离经叛道的细胞。因为突变会随着时间累积,状态最为异常的细胞通常会在雪花的顶端被发现。但他们在有机会成为叛徒细胞之前,就会分裂形成新的菌落。
这种机制也能使群体特性在酵母中进化。每个雪花分支释放出的细胞产生的突变都会被传递给下一个菌落中的所有细胞。因此,随后的雪花开始将新的群体特征――比如细胞的大小和数量,或者自杀细胞出现的频率和位置――作为进一步进化的“原材料”。从那时起,需要适配的是组合方式,而不是单个细胞。
有关酵母的研究结果并非侥幸成功。2014年,拉特克利夫和他的同事为了获得更大的细胞对单细胞藻类衣藻进行了同样的筛选,并再次发现了大量的菌落。为了应对人们对于他的人工选择技术人为干涉过多的批评,他和赫伦使用了一种更为自然的选择压力重复了衣藻实验:他们引入了一群以衣藻为食、并倾向于选择较小细胞的草履虫。一种多细胞生物又一次快速出现了:在750代之内――大约1年时间――5组实验群体中的两组开始形成多细胞生物并以群体的形式进行繁殖,研究小组在1月12日的bioRxiv预印本中写道。
如果多细胞生物的出现如此的简单,为什么复杂的生物体要在生命起源后的数十亿年后才被牢固地建立起来呢?传统上,研究人员将此归咎于早期大气的低氧水平:为了获得足够的氧气,生物体需要尽可能高的体表面积与体积比,这就迫使它们保持着较小的体积。只有在大约10亿年前氧气水平上升之后,才能产生更大的多细胞生物。
然而,2015年,英国剑桥大学的古生物学家尼古拉斯·巴特菲尔德(Nicholas Butterfield)提出,低氧水平实际上有利于古代海洋生物向多细胞生物的进化。体积更大的多细胞生物――有多个鞭毛――更擅长将水从细胞膜上扫过以获取氧气。古代海洋中匮乏的营养物质将有助于推进下一个阶段的进化,即特殊细胞类型的进化,因为更复杂的生物体可以更高效地获取食物。至于为什么复杂的生物需要这么长时间才能出现,巴特菲尔德认为这种滞后反映了进化出多细胞生物更为复杂的基因调控机制所需的时间。
巴特菲尔德的理论是“非常优雅和简单的,建立在物理和化学的基本原则之上,整合入深层的地球化学、生物地球化学和生物物理环境。”加州大学戴维斯分校的进化生物学家理查德·格罗斯伯格(Richard Grosberg)说。
一旦生物跨越了多细胞性这一门槛,就几乎开弓没有回头箭了。在许多遗传谱系中,细胞和器官的种类数量持续增长,并且发展出更为复杂的方式来协调它们的活动。拉特克利夫和瑞典于默奥大学的理论生物学家埃里克·利比(EricLibby)4年前提出,棘轮效应占据了主导地位,推动了多细胞生物在复杂性方面势不可挡的增长(《科学》,2014年10月24日,第426页)。复杂生物体的细胞越专门化、越相互依赖,就越难恢复单细胞的生活方式。英国牛津大学的进化生物学家盖伊·库珀(Guy Cooper)和斯图尔特·韦斯特(Stuart West)最近在数学模拟图像中证实了这一说法。5月28日,库珀和韦斯特在《自然生态与进化》杂志上写道,在形成更为复杂的生物体时,“劳动分工不是结果,而是驱动因素。”
从一个细胞到许多细胞的最初转变触发了一轮不断增长的生物复杂性,如今丰富多彩的多细胞生命就是结果。
资料来源 Science
责任编辑 彦隐