细胞是生命结构与功能的基本单位。生命科学发展到今天,人类已有可能鉴定出完成细胞功能所需的全部基因。但有关的基因是如何作用以产生细胞组织的?细胞内的所有基因是按什么规则协同作用的?进一步弄清楚这些问题,将有助于我们充分了解细胞,从而充分了解生命本身——

细胞学说是生物学史上最重要的理念之一;它提出,所有的生命形态都是由细胞组成的。细胞是显示生命特征各项功能最简单的单位;细胞生物学已帮助我们揭示了种类极多的生命形态是如何组织起来并顺利运转的。本文回顾细胞作为生命结构与功能的基本单位的发现历程,特别聚焦于最终导致生长、发育与进化的细胞增殖的机制与调控,并对有助于进一步阐明细胞功能的未来发现做一些推测。
寻找生命的基本单位
在科学上,通常是技术导致发现。显微镜的发明导致细胞的发现。17世纪早期,在荷兰通过改进透镜,造出了用于研究昆虫和其他小生物的简单显微镜。英国科学家罗伯特 · 虎胡(Robert Hooke)应用复式显微镜观察软木塞的切片,在他1665年出版的《Micrographia》一书中,他作图描述了他之所见,并将有壁的小室命名为“细胞”。
不久,尼海米阿 · 格鲁(Nehemiah Grew)与马塞罗 · 莫尔皮奇(Marcello Malpighi)分别作图描绘了植物细胞的全面特征,形成了“植物组织大都由聚集成团的细胞组成”的观点。又不久,莫尔皮奇、安东 · 凡 · 列文虎克(Anton Van Leeuwenhoek)和简 · 施旺达姆(Jan Swammerdam)等人成为在动物体上认出细胞的首批科学家。他们描述了血液里的血球;但那时还没人提出动物的固体组织也是细胞组成的。列文虎克还发现了生长在植物提取液中的他称作微动物的单细胞生物。显微镜观察的改进,导致对细胞的更细致的描述。1766年,亚伯拉罕 · 特伦布利(Abraham Trembley)报道了对微动物(原生动物synhedra)的繁殖的观察;这可能是对细胞二均分裂的首次报道。
物质的原子属性被逐渐接受,也帮助了细胞学说的形成。到18世纪,当生物学家解释在显微镜下看到的植物和动物的结构时,考虑生命物质的基本亚单位已经不足为奇了。关键的臆测,是劳仑兹 · 奥肯(Lorenz Oken)于1805年提出的。他曾论证,多细胞的植物和动物是微小的、有生命的“浸液虫类”(诸如生长于动物和植物提取液的原形动物)的集合体。
嗣后,植物学家马修斯 · 施莱登(Mattias Schleiden)与动物学家西奥多 · 施旺(Theodor Schwann)注意到细胞间的一些共同特征,加以归纳、概括,形成了细胞学说。他们在1839年写道,“我们已观察到,所有的生物体是由本质上相似的部分——细胞——组成的。鲁道夫 · 弗乔瓦(Rudolf Virchow)在其1858年出版的《细胞病理学》一书中写道:“每个动物作为生命单位之总和而出现,每个单位显示全部的生命特征。”这个发现——细胞是所有生物体的结构与功能的基本单位——是生物学历史上的一座里程碑。
细胞显示全部生命特征
施莱登与施旺虽然正确地阐明了细胞学说,但他们还曾对细胞的形成困惑不解,认为它们产生的过程类似沉淀与结晶。其他学者,特别是研究植物细胞的巴瑟莱米 · 杜莫狄尔(Bathelemy Dumortier)与研究动物细胞的罗伯特 · 雷马克(Robert Remark)则认识到,上一代细胞通过二均分裂,产生下一代细胞。这个观点,为弗乔瓦所大力支持,他的名句“细胞来自细胞”广为传播。
研究深入到细胞增殖这一步,细胞的惊人的复杂性及其机器似的特点开始被揭示。细胞增殖的核心,是最终解释了孟德尔遗传定律的藉分子机制运作的遗传系统。19世纪80年代,孟德尔得出结论认为,植物的性状(诸如种子形状、植株高度)决定于成对的因子,成对因子中的一方来自雄性植物,另一方来自雌性植物。他认为,在杂交植物中,这些因子保持独立,并在进一步杂交中作为独立实体而传递。只是到了20世纪初,研究人员发现孟德尔所谓的因子的行为反映了染色体在细胞分裂中的行为,孟德尔的上述结论才得到更科学的认可。
大多数细胞含有单个细胞核,它们在有丝分裂与细胞分裂时增殖。18世纪80年代,沃尔瑟 · 弗莱明(Walther Flemming)与爱德华 · 斯特兰斯伯格(Edward Strasburger)观察到,在有丝分裂进程中,伸长了的染色体纵向分裂,然后缩短、变粗。纵向的各半分开,进入子代细胞核。与此观察同时,爱德沃德V · 贝尼(Edouard Van Beneden)证实,线虫受精卵内的染色体来源于卵子与精子中的等数染色体。这导致奥格斯特 · 魏斯曼(August Weisman)提出,遗传系统是以染色体为基础的。
科学家们在形成有关细胞增殖机制的初始理念的同时,力图搞清细胞增殖与生物体生长发育间的联系。单细胞微生物的群体生长需要细胞分裂;到19世纪后叶,研究人员认识到,多细胞生物的生长,也是基于细胞分裂。19世纪60年代,鲁道夫 · 考立柯(Rudolf Kölliker)观察到,早期胚胎的卵裂反映一系列的细胞分裂,所生成的细胞随即分化为特种组织。到19世纪80年代,科学家们得出结论:所有多细胞生物,不论其如何复杂都来自单细胞。
细胞增殖对生长发育的极端重要性可直接感到;细胞死亡也是生长发育的关键则没有那样的直觉性。但在19世纪,支持后一观点的资料逐渐增多。病理学家观察了健康组织中走向死亡的细胞。描述这一现象的新词“细胞凋亡”(“apopsis”)是1972年新造的。正是细胞凋亡在线虫发育上的作用的发现,肯定了细胞凋亡的积极功能。线虫卵进行高度特化的细胞分裂,形成由固定总数的细胞组成的成虫。对细胞世系的监测揭示,在发育的一些特定阶段,某些细胞经常进行凋亡;而且分离出了细胞凋亡被抑制的突变体,它们死亡时具有更多的细胞。这些试验提示,细胞凋亡是有程序的。在特定发育阶段,在某些细胞,细胞凋亡径路开通,一系列蛋白质酶被激活,导致这些细胞崩解。近年来,研究人员已鉴定出许多启动固定的细胞死亡程序的基因。
没有为更大的利益而自杀的细胞很容易误入歧途。19世纪中叶,雷马克与弗乔瓦认为,有病组织的细胞来自正常组织,意谓功能不良的细胞导致疾病。这一认识在医学上有很大的重要性,因为它把注意力集中到细胞行为(最终是分子行为)的改变,把它作为理解疾病的最重要因素。
对癌症的研究最能说明这个问题。早期的病理学家曾认为,癌起因于细胞增殖失控的发育不正常的组织。到20世纪70年代,研究人员已证明,遗传变异(诸如染色体重组、某些病毒基因之存在)导致细胞癌变。在小鸡Rous肉瘤病毒里鉴定出src癌基因,在哺乳动物鉴定出相应的癌基因;这些都是重要的发现。随后又发现了许多被激活时能促进细胞生长和分裂的其他癌基因。
一些抑癌基因也被鉴定出来了,它们在正常情况下抑制细胞的生长和分裂,当它们失活时,其所在细胞就要癌变。其他与癌有关的基因,包括监察基因损伤并激活细胞凋亡径路的一些基因,则参与安全巡察,如果个别细胞的基因损伤很大,可能诱发癌变的话。编码P53的基因是人体肿瘤中最常见的突变基因,已以“基因组监护者”著称,因为它是正常监视基因组损伤所必需。缺少P53的细胞,即使受到广泛的基因损伤,仍然存活。癌细胞也能是因细胞凋亡径路有缺陷;在这些细胞中不能激活凋亡径路,所以基因损伤的细胞仍存活。如果这些存活细胞中控制生长和分裂的基因受到损伤,它们将选择性地增殖,并在原来的组织中占优势。最终,它们就转移并扩散至身体的其他部分。
细胞周期循环往复
细胞增殖是遗传、生长、发育的基础,因此细胞里所有的基因需要复制并分开,以便整个基因组能准确地传递到每个子代细胞。这一过程是靠称作细胞周期的一系列事件来实现的。
细胞周期中基因组复制与分开所需的两大主要事件是S-期(DNA合成期)与M-期(丝裂期)。在S-期,细胞复制其染色体DNA。这个过程起先藉细胞周期的标记研究及精确测定细胞周期中细胞DNA含量的改变来鉴定。这些试验显示,DNA合成只发生在细胞周期早期的有限间期。在M-期(发生于细胞周期末期),复制好的染色体分开并进入由细胞分裂形成的两个子细胞。子细胞形成与S-期之间的间期,以及S期结束与M期之间的间期,分别叫做G1期与G2期(G代表间断)。这样,细胞周期就可分成4个期:G1期、S期、G2期和M期。
这些就是运行细胞周期的分子机器的工场价值。想想DNA复制吧。关于这个过程,没有什么比1953年对DNA结构的发现揭示得更多的了。该发现解决了并存的两个问题,即DNA怎么精确地复制,以及怎么给信息编码?DNA的碱基序列是这样给信息编码的:双股碱基对结构的解开,产生一对单股模板,用来合成精确的互补股。
掌握了DNA的结构,搜寻细胞复制背后的机器就比较容易了。DNA多聚酶的发现,提供了第一把钥匙。该酶能给先已存在的DNA模板合成一条互补股。接着,许多其他有关的酶被相继发现,其中有局部异构酶和解旋酶,它们是解开DNA股的;引物酶和DNA多聚酶,它们是合成新的DNA股的;还有把DNA股拼接起来的连接酶。在细胞的S期,这些酶集合于DNA的称作“复制起点”的特化区域,DNA复制从这儿向两个方向展开。复制的两条逐渐分开并形成DNA“泡”,最终完成复制过程。
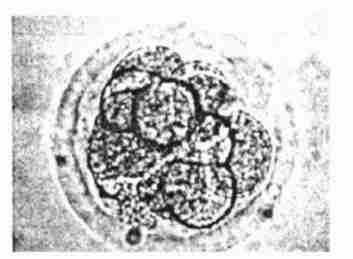
显微镜下的细胞分裂图
像任何复杂的机器一样,复制机器也需要调控。例如,复制得以开始之前,一些启动因子必须连结到复制的起点。这些因子包括起点识别复合物作为启动蛋白质的“着陆垫”。这些启动蛋白质随后负载DNA复制所需的一些其他蛋白质。正常情况下,在每一细胞周期,这个过程只被激活一次,所以只有一次S-期发生。否则,细胞会一遍又一遍地多次复制其染色体。
准确的DNA复制,是细胞周期增殖机制中的首要内容。其次是首次描述于19世纪的M-期的有丝分裂。有丝分裂开始时,紧缩了的复制的染色体(每个由纵向并列的成对姊妹染色线组成)逐渐显现。染色体在细胞中央排列,并与纺锤丝连结。在细胞相对的两端,各有一叫做中心体的结构,纺锤丝在这两者之间延伸。然后,染色线互相分开,各自向细胞的一边移动,它们在那里集合成两个细胞核,进入新生成的子细胞。
对这一机制特别重要的,是组成纺锤丝微管的微管蛋白多聚体。这些微管显示动力学上的不稳定性——变动于生长与萎缩之间。中心体使微管固定,因此也使纺锤丝固定。从中心体发射出的其他不断生长的微管“探查”细胞内部,直至其游离端与动粒、连接而被固定,动粒是每条染色线上着丝粒上的结构。这个过程使每条染色线与一个中心体相连接。
只有当姊妹染色线上的动粒分别与细胞上的两个中心体连接起来,才发生稳固的连接。这时,每对染色线中的各半分别向细胞相对的两端移动,姊妹染色线间的吸附力消失。这个分开的动作由微管中收缩蛋白与动力蛋白的联合动作驱使微管相对滑动而实现。这个过程,把分子水平上的DNA复制与细胞水平上的染色体分开联结起来,并确保基因组的精确的复制与分离。
(待续)
[Science,2000年9月8日]