自麦克斯韦引入著名的“妖”概念以来,信息与物理学之间的联系就已经不言而喻。如今,信息甚至可能成为连接物理学与生物学的关键概念。

在物理学家看来,生命就像魔术。生物取得的成就如此耀眼又如此神秘,我们甚至因此忘记了生物也是由寻常原子构成。然而,如果生命的奥秘并不是构成它的那些物质,会是什么呢?是什么赋予了生物体如此不同的生命活力,使它们如此与众不同?这正是埃尔温 • 薛定谔(Erwin Schrödinger)1943年在爱尔兰都柏林所做的一系列著名讲座上提出的问题。第二年,他又在颇有影响力的著作《生命是什么?》中正式发表了这个问题。
薛定谔是理论物理学巨匠,也是量子力学的创始人之一,而量子力学是迄今为止最成功的科学理论。例如,将量子力学应用于电磁场时,它能正确预言电子的异常磁矩,精度超过10位有效数字。量子力学几乎一下子就解释了所有非生命物质的性质,从亚原子粒子到原子、分子,再到恒星,似乎就没有它不能解释的。然而,量子力学没能解释生命物质,这令人沮丧。另外,虽然生物学在量子力学诞生之后的几十年里进展神速,但生命之谜仍旧没有揭开。没有人能肯定地回答“生命是什么”以及“生命如何开始”。
在被问及“物理学是否能解释生命”时,大多数物理学家的回答是肯定。然而,更为切题的问法应该是:目前已知的物理学是否能够解释生命,或者说,我们是否需要全新的基础物理理论?20世纪30年代,量子力学的众多设计师——最为知名的当属尼尔斯 • 玻尔(Niels Bohr)、尤金 • 维格纳(Eugene Wigner)和维尔纳 • 海森堡(Werner Heisenberg)——就已经预感到:在有关生命物质的物理学中,的确存在一些全新和不同的东西。
对此,薛定谔并没有明确表态,但他对这种可能性持开放态度。“我们可能会找到在生命物质中普适的全新物理学定律,必须做好准备。”他推断说。不过,薛定谔并没有明确提出这类物理学定律究竟是什么。
这类问题已经超越了纯粹的学术问题。天体生物学的核心目标就是在地球之外寻找生命的踪迹,但如果没有生命的定义,那就很难精确知道在寻找什么。举个例子,美国宇航局正在筹划一次飞行任务,目标是让探测器穿过土卫二恩科拉多斯(我们已经确定这颗土星卫星上存在有机分子)冰壳裂隙中喷射出的羽状物。那么,我们要怎么才能让怀疑论者相信这些羽状物中包含生命,或者包含曾经是生命的有机体的碎屑,而不是某种形式的前生命物质?与测量磁场这样的行动不同,对于生命,科学家暂时还没有任何形式的标准,以量化化学混合物向已知生命形式演化的过程,更不用提外星生命形式了。
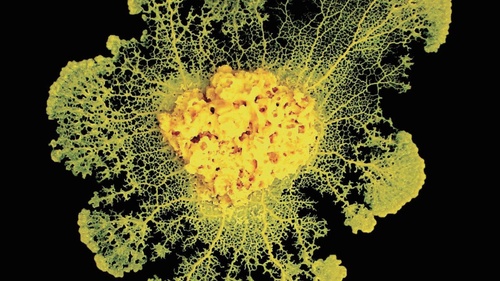
图1 黏液菌。这些自由单细胞构成的集合有时会结成合作联盟,表现得就像拥有共同计划的单一有机体一样
大多数天体生物学家重点关注我们所知的生命特征。例如,美国宇航局在20世纪70年代开展的“海盗”系列探测火星的任务,就利用了可食用的陆地生物的营养物质汤搜寻火星上碳代谢的踪迹。另一种经常被天体生物学家提及的生命特征就是同手性。虽然物理学定律并不关心左右手性的问题,但目前已知的地球生命确实都使用了左旋氨基酸和右旋核糖。然而,无机土壤中的化学环境也能产生类似新陈代谢的效果,且没有生命参与的化学循环在重复多次后也足以产生同手性现象。因此,这些天体生物学家设想的生命特征并不是决定性的,要想确认地球之外的生命就更难了。
天体生物学家把希望寄托在探测太阳系外行星大气层中的氧上,但这也并不是可靠的选择,大气层中的氧并不是光合作用的明确标志,因为非生物过程也能产生含氧大气层。我们缺乏独立于生化基底的关于“活着”的定义。是否存在更深层的普适原理,为我们指出毫无争议的生物标志,即便是我们目前尚不知晓的生命形式也能凭此加以判断?
两种文化
物理学与生物学之间的分歧可不只是复杂性上的问题,这两门学科在概念框架上也存在本质上的不同。
物理学家通过能量、熵、分子力、反应率这样的概念研究生物。生物学家则完全不同,他们考察的是诸如信号、编码、转录、转换这样的概念——也是信息的语言。令人惊奇的全新基因编辑技术CRISPR就是很好的例子。这项技术赋予了科学家编辑生命密码本的能力。欣欣向荣的生物物理学试图通过对各类生物控制网络的信息流动和存储的模式建模以跨越这两门学科之间的概念鸿沟。
生命在各个层面上(不只是DNA)都会进行信息存储和处理。基因——起到加密指令集作用的DNA序列——可以通过化学信使开关其他基因,并且常常会形成复杂网络。那些化学回路就像是电子元件或计算组件,有时也会充当执行逻辑运算的模块或门。
在细胞层面上,有多种物理机制会发出信号,进而促成合作行为。图1所示的黏液菌就是极好的例子。它们其实只是诸多单细胞构成的集合,但可以自组织形成引人瞩目的形状,有时还会表现出统一的行为,就好像它们是单个有机体一样。类似,像蚂蚁和蜜蜂这样的社会性昆虫会互相交换复杂信息并且参与集体决策,而人类大脑则是极为复杂的信息处理系统。
生命的信息基础让部分科学家发布了这样的非正式声明:生命=物质+信息。然而,要想通过这个等式做出真正有效的解释和预测,就必须构建能够把物质和信息联系起来的理论框架。这种联系的第一条线索出现于1867年。当时,在写给朋友的一封信中,苏格兰物理学家麦克斯韦构想了一种能够感知盒中气体分子的微小生物。这种生物很快就顶着“麦克斯韦妖”的名头为大家所知。如图2盒中场景所示,这个小东西能通过操作隔板及小门将所有运动速度较快的分子赶到盒子左侧,把运动速度较慢的分子赶到盒子右侧。
麦克斯韦妖

图2“麦克斯韦妖”示意图
图2展示的是一个装有气体的盒子,中间有一块带着小门的隔板把盒子分成两个小房间,气体分子(灰色圆)可以一个一个穿过小门。控制这扇小门的就是麦克斯韦1867年的脑力劳动产物:一只时刻观察着随机运动分子的小妖,它会尽职操作小门以保证运动速度较快的分子从右侧房间运动到左侧,而运动速度较慢的分子则被它“赶”到右侧。于是,我们就可以利用这样一种机制把无序的分子运动转变成有方向的机械运动。
在其诞生之后的几十年里,麦克斯韦妖的概念一直是人们不愿直面的物理学核心问题,大多数人都只是把它当作理论谜团。麦克斯韦提出这个思想实验一个世纪后,一只真正的妖在他的出生地爱丁堡的一间实验室中诞生了。这个实验由一个套在杆上、可以前后滑动的分子环构成,杆的末端装有塞子,中部待着另一个分子。这个分子有两种状态:一是阻隔分子环,使其不得前进;二是放行,让分子环通过。因此,这个分子就起到了门的作用,与麦克斯韦最初构想的小门类似。
自那之后就兴起了一个制造这种“妖”设备的产业,其中包括芬兰阿尔托大学尤卡 • 佩科拉(Jukka Pekola)领导的纳米科学研究小组和石溪大学德米特里 • 艾弗林(Dmitri Averin)共同研制的信息驱动冰箱。在这款冰箱中,充当气体分子的是一枚处于双面纳米盒中的电子,且这个纳米盒与一个热浴耦合。冷却循环利用纳米盒的两种简并态获取一定电子能量。循环开始时,电子处于一种确定的非简并态。外部电场可以把电子能量提高到简并能级,在这种能量下,电子可以处于两种状态中的任何一种,且概率相等。
这种不确定性的介入代表电子的熵增加,而热浴的熵及温度则相应减少。这个时候,妖——第一个电子盒耦合的另一个单电子盒——就开始发挥作用。它会侦测到电子究竟处于哪种状态,并且自动把这个信息反馈给驱动场,后者则根据这条信息迅速将电子的状态调回初始时的非简并态并完成整个冷却循环。研究人员发现,每次循环都会产生一个比特的信息——表明电子处于循环之中——这个过程能以75%的平均效率从热浴中汲取热量。麦克斯韦没错,信息真的可以充当某种形式的燃料。
因为分子运动速度是对温度的一种度量,所以从效果上来说,这个小妖会利用有关分子的信息在盒内创造一个热量梯度。然后,工程师就可以从这个梯度中汲取能量并加以利用。从表面上看,麦克斯韦无视热力学第二定律,设计了一部由信息驱动的永动机。
为了解决这个悖论,我们就必须量化信息并将其正式整合到热力学定律中。20世纪40年代末,克劳德 • 香农(Claude Shannon)构建了现代信息理论的基础。他将信息定义为不确定性的下降。举一个统计掷硬币结果的例子,用我们熟悉的二进制数字,或者说比特,来表征掷硬币得到的信息——正面向上还是背面向上。综合应用香农的信息理论和热力学定律,我们就会把信息认定为负熵。小妖为提升热力学有序度获取的所有信息都必须以某些阶段的熵增为代价。例如,为了让小妖重启循环,就必须在适当的时候擦除并重置它的记忆库。
麦克斯韦只是把他的妖当成思想实验,但如今纳米技术的发展让我们得以在实验层面上实现这个想法(比如前文所述的电子盒)。不过,生命在几十亿年的发展历程中一直在创造并利用各种妖。我们的身体中到处都是它们的身影——复制DNA,沿着纤维运送“货物”,把质子抽送到细胞膜中的各类分子机制等,运作方式非常接近理想热力学极限。它们利用热力学第二定律获取能量。人类大脑也在回路中使用了一类妖——电压门控离子通道——以传送电信号。有了那些离子通道,大脑才能用相当于一只昏暗电灯泡的能量完成工作,且计算能力相当于一台兆瓦级超级计算机。
生物信息的环境属性
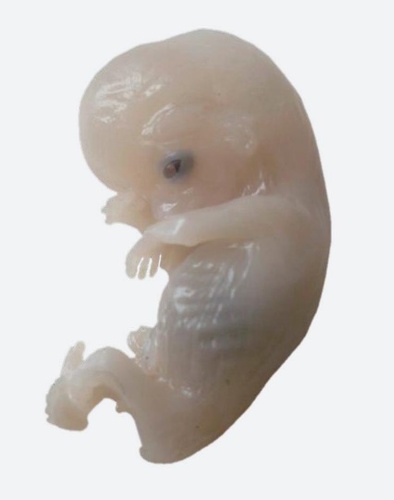
图3 8 ~ 9周的人类胚胎,38毫米长
这种“妖”机制只是生命信息的冰山一角。生物信息的作用远不止优化能量分配,它还常常起到管理者的作用。想想受精卵发育成胚胎(图3)的方式。发育过程的每个阶段都受到信息网络的监督,而这些信息网络在进化过程中已经微调适应了物理过程和化学过程,所有这些安排的目的,都是为了让胚胎最终能以正确的结构和形态出现。
利用基因调控网络中的信息流模拟胚胎形成过程的尝试已经取得巨大成功。从化学角度上说,加州理工大学的埃里克 • 戴维森(Eric Davidson)和他的合作者已经绘制了管控海胆早期发育过程的基因网络的完整线路图。通过追踪信息流,这个研究小组编写程序来模拟基因网络演化过程。他们在每个阶段都会将回路状态的计算机模型同实际观察到的海胆发育阶段作比较,结果惊人地吻合。研究人员还在计算机模型中考虑了化学沉默特异性基因的影响,以预测突变胚胎的情况。结果,模型预测再一次与实验观测相符。
普林斯顿大学托马斯 • 格雷格(Thomas Gregor)和威廉 • 比亚莱克(William Bialek)领导的研究小组一直在研究果蝇发育的初期阶段,尤其是果蝇的标志性形态特征最初是怎么出现的。在发育过程中,细胞需要知道三维空间中它们与其他细胞之间的相对位置。那么,它们是怎么获得那些位置信息的呢?很早之前,我们就知道细胞可以充当某种以化学梯度为基础的GPS,而这些化学梯度又由特定基因的表达水平管控。普林斯顿大学的这个研究小组最近把重点放在了四个所谓的“间隙基因”上。这些基因通过在身体平面图中创造间隙或条带为胚胎基本模型打下基础。研究人员发现,细胞利用贝叶斯概率从基因表达水平中提取最佳位置信息,并且误差小到了令人咋舌的1%。他们将一个优化贝叶斯模型应用到诱变菌株上,并且成功预测了修正后的菌株形态。
所有这些实验分析都提出了一个至关重要的哲学问题,直指物理学与生物学概念不匹配问题的核心。我们现在通常把针对基因调控网络和贝叶斯算法应用的研究看作现象学模型,在该模型中,“信息”是生成真实有机体的逼真模拟的简便替代或标签。然而,麦克斯韦妖的思想实验告诉我们,信息其实是一种可以深刻影响物质行为的物理量。按照香农定义,信息不只是一种非正式参数,更是一种在热力学定律中有明确位置的基础物理学变量。
香农强调,他的信息理论纯粹处理的是信息流的效率和能力,不涉及任何信息传播的意义。然而,在生物学中,意义或语境非常重要。我们要怎样才能从数学角度捕捉指令信息、监管信息或者语境信息的属性呢?一种方法是分子生物学的所谓“中心法则”(克里克和沃森提出DNA双螺旋结构之后10年左右杜撰的术语),即认为信息只会单向流动,从DNA流向蛋白质再流向有机体。我们也可以称这是一种“自下而上”的流动。
现在,我们已经知晓,生物学中的信息转移其实是一种双向过程,涉及反馈循环和自上而下的信息流。例如,如果培养皿中培养的细胞太过拥挤,它们就会停止分裂,这个现象叫做接触抑制。另外,在国际空间站开展的微生物实验则表明,零重力环境下的细菌基因表达与地球环境不同。显然,系统层面上的物理力能够影响分子层面上的基因表达。
塔夫斯大学艾伦发现中心迈克尔 • 列文(Michael Levin)及其同事的工作就是自上而下信息流的极好例子。列文领导的这个小组正在研究在某些生物的生长和形态控制实验中,为什么系统层面的电模式与机械力及化学模式同等重要?
健康细胞是电极化的,它们通过抽送离子在细胞壁间维持了上百毫伏的电势差。相较之下,癌细胞是去极化的。
列文研究小组发现,在多细胞生物中,各组织之间的细胞极化模式在生物的生长、发育、伤口愈合以及器官再生过程中都发挥了关键作用。研究小组运用化学手段打乱了这些生物电模式,成功催生了全新的生物形态。某种扁涡虫为开展这类实验提供了便利(图4)。在正常情况下,如果把这种扁涡虫砍为两截,原先的头部就会长出新尾巴,原先的尾巴则会长出新脑袋,这样就产生了两条完整的扁涡虫。不过,在改变伤口附近的电极化状态之后,我们得到的就不是两只正常扁涡虫了,而是两只双头扁涡虫或双尾扁涡虫。
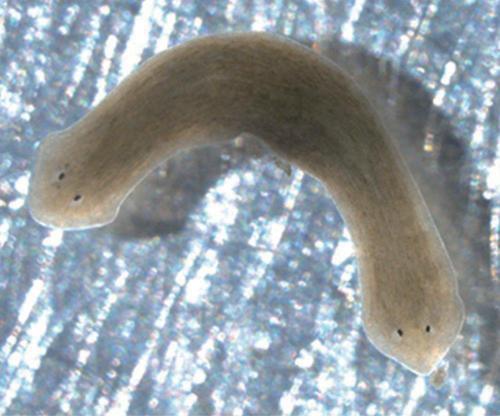
图4 人为操控电极化状态产生的一只双头扁涡虫
令人惊异的是,如果我们继续把双头扁涡虫截为两段,它们并不会回到正常的表现型,即并不会变成正常的单头扁涡虫。双头扁涡虫在一分为二后产生的仍是双头扁涡虫,双尾扁涡虫也是如此。虽然双头/双尾扁涡虫与正常扁涡虫的DNA完全一致,但在外观上似乎就成了两个物种。系统的形态信息必然是以分散的形式储存在截断的组织中,并且各自在基因层面上指导正确的再生过程。不过,这个过程到底是怎么发生的呢?电信号密码是否与遗传密码协同工作?
表观遗传学是指一些超越基因层面且能决定表现型的因素,比如总物理力。我们现在对表观遗传过程中的信息存储、加工和传播机制几乎一无所知,但它们在生物学中起到了至关重要的作用。要想在这方面取得进展,我们需要研究不同类型的信息模式(生物电信息、化学信息、遗传学信息)之间如何发生相互作用,以产生能够管理生命物质组织并将其翻译为特定表现型的调控网络。
从信息角度而非分子角度思考生命物质涉及的物理学问题与计算科学中软件与硬件的差别类似。全面理解某个特定的计算机应用不但需要掌握软件工程原理,而且对计算机电路涉及的物理学也需要有同样的认识。类似,只有在我们全面了解生物信息机制之后,才能真正理解生命。
动力学的新概念
自牛顿时代以来,主导物理学的一直是基本二元论。虽然物理状态会随时间而改变,但基本物理学定律通常是永恒不变的。这一假设是汉密尔顿动力学、轨迹可积性和遍历性的基础。然而,把物理学定律不会改变这个假设应用到生物系统中时,就有点“水土不服”了。在生物系统中,信息的动力学模式与时间依赖的化学网络相耦合,并且信息表达——比如基因的开启——既可以受整体或系统物理力控制,也可以由局部化学信号管控。
以高度多样性、创新性和不可预测性为特征的生物演化过程与非生命系统的时变方式形成了鲜明对比。然而,生物学并不等同于混乱,我们也能在这个领域中找到许多能确实发挥作用的规律。以具有普遍性的遗传密码为例,核苷酸三联体CGT会编码精氨酸。虽然这个现象或规则目前还没有出现过例外情况,但如果你把它看作自然定律(就像绝对正确的万有引力定律一样),那绝对是一种错误。几乎可以肯定的是,CGT编码精氨酸这种现象必然是上百万年前从某些更早、更简单的规则演化而来。生物学中还有许多这样的案例。更有现实意义的生物系统变化描述应该是把动力学规则的变化视为系统状态的函数。这种随状态改变的动力学机制(状态依赖动力学机制)开辟了包含无数新奇行为的广阔天地,但远不是正式的数学理论。那么,这种新动力学机制与正式数学理论之间还差点什么呢?我们拿国际象棋来作类比。在标准国际象棋中,游戏系统封闭且规则固定。从正常初始状态开始,国际象棋棋手可以自由探索棋局的后续状态。虽然后续的可能状态有很多,但会受到不可改变的对弈规则的限制,因而也只会是所有可能出现的棋局的微小子集。也就是说,虽然规则容许的棋局状态有许多,但规则禁止的棋局状态更多,比如,所有象都处在相同颜色的方格内就是规则禁止出现的一种棋局状态。
现在,我们想象一种修正版的国际象棋:规则可以随棋局的整体状态而改变,即一种系统层面、自上而下的标准。举一个简单例子,如果白方即将获胜,那么规则就会变为黑方的兵可以前后移动,而非只能向前。在这种“进阶版”国际象棋中,整个系统处于开放状态,出现的棋局状态也会是固定规则下的标准国际象棋所禁止的。生物学其实就和这种想象中的国际象棋类似,其中,生物体同样也是开放系统,容许非生命系统禁止的事情发生。
为了探索把这种状态依赖动力学机制放在以自上而下信息流为特征的简单模型中会产生什么效果,我在亚利桑那州立大学的研究小组运用了一种修正版的一维细胞自动机(cellular automaton)。标准细胞自动机应该是一排细胞——用方格或者像素来模拟——要么是空的,要么是实的(分别用白和黑表示,当然也可以用其他方法表示)。然后就要用固定规则根据每个细胞及其最近邻居的现时状态来更新它们的下一步状态。整个系统有256种可能的更新规则。
这个细胞自动机游戏是这么玩的:先选一个初始细胞模式——用二进制序列(0或1)表示会比较方便——然后再反复应用挑选出来的更新规则驱动系统演化。许多更新规则只会引出毫无意义的结果,但有一些会产生具有相当复杂性的精致模式。为了应用机制随状态改变的修正版细胞自动机,我的同事阿利萨 • 亚当斯(Alyssa Adams)和萨拉 • 沃克(Sara Walker)通过计算机手段耦合了两种标准细胞自动机,一种代表生物体,另一种代表环境。
然后,他俩允许生物体的更新规则在每次更新换代时改变。那么,每次更新时如何在256条规则中挑选呢?为此,他们把生物体细胞自动机中的细胞捆绑成相邻“三胞胎”形式——000、010、110,以此类推;然后再将每个三胞胎的相对出现频率同环境细胞自动机中的相同模式作对比。这样一种设定就把更新规则改变成了生物体状态和环境状态共同的函数。更新规则是生物体状态的函数,意味着它有自我参照;是环境状态的函数,意味着它是开放系统。
亚当斯和沃克在计算机上运行产生了上万个研究案例,并试图从中找到一些有意思的模式。他们想要认证那些既具有开放性(生物体不会迅速通过循环返回初始状态)又具有创新性的进化行为。这里的创新性是指:无论从何种初始状态开始,观测到的生物体状态序列都永远不会与256种固定规则下的细胞自动机产生的状态模式相同。这就好比是在我们前面提到的修正版国际象棋中出现了双方4个象最后都处在相同颜色方格中的对局状态。虽然这个系统已经如此开放,但这种创新性进化行为仍十分稀少,只出现了少数明确的案例。计算机对细胞自动机的模拟花了很长时间,但亚当斯和沃克的发现已经足够证明,即便在他们的这个简单模型中,状态依赖动力学机制也能产生通往复杂性和多样性的新进化道路。他们的工作还证明,只是加工处理信息是不够的。要想全面体现生物学多样性,约束信息处理过程的规则本身也必须进化。
量子边缘的生命
如果生物学应用我们目前尚不了解的全新物理学,比如前面所说的状态依赖动力学机制,那么这种物理学会在简单分子与生物细胞之间的哪个节点上出现?细胞自动机模型确实能为我们提供许多信息,但它们只是计算机勾勒出来的“动画”,并不是物理学,也没有告诉我们任何有关去哪儿寻找这些新兴现象的信息。实际上,标准物理学中已经出现了状态依赖动力学机制,而且我们也相当熟悉,那就是量子力学。
用相干波函数描述处于孤立状态的纯量子态,我们就可以根据充分掌握了的数学方法(即“幺正演化”)准确预测这种量子态的演化过程。不过,在测量之后,这个量子态就会突然改变,这个现象叫作波函数坍缩。如果测量结果理想,那么量子态的跃迁会把系统投射到与被测量的可测量对应的可能本征态上。这时候,幺正演化就被玻恩规则代替,后者能预测各种可能测量结果出现的相对概率,并且将非决定性或不确定性引入量子力学。这标志着从量子领域到传统领域的转变。那么,量子力学能告诉我们是什么让生命生生不息吗?
薛定谔在他著名的都柏林系列讲座中提出,量子力学应该能解释遗传信息储存的稳定性问题。早在克里克和沃森阐述DNA结构之前,薛定谔就已经推断出,这类信息在分子层面上必须储存在他命名为“非周期晶体”的结构中,事实证明,这个说法正是对核酸聚合体的感性描述。然而,量子现象是否可能在生物体中起到更为普遍的作用,这个问题仍旧没有定论。
从薛定谔都柏林讲座到DNA结构提出的几十年里,大家普遍秉持的主流观点是:在生命物质这种温暖、嘈杂的环境中,量子现象会被扼杀,经典化学就足以解释生命。不过,最近的十多年里,大家开始觉得非平凡量子现象(比如叠加、纠缠和隧道效应)终究可能对生命颇为重要,对这些现象与生命间联系的兴趣也因此与日俱增。虽然怀疑的声音仍旧不小,但量子生物学这个新兴领域现在已经进入深入研究阶段,且重点研究课题涵盖范围甚广,比如光合作用中相干能量的传输、鸟类磁场罗盘以及苍蝇的嗅觉反应。
在纳米尺度上研究生命物质的量子特性是一项非常困难的挑战。对生命运行至关重要的那些系统可能几乎不涉及任何自由度,大幅偏离热力学平衡态,并且与热环境强烈耦合。然而,事实就是如此,非平衡量子统计力学领域的确是有可能产生全新物理学的地方。
一组可能与此相关的实验是通过有机分子测量电子电导性。最近,伽柏 • 瓦泰(Gábor Vattay)及其同事宣称,许多在生物学上颇为重要的分子,比如蔗糖和维生素D3,都拥有独特的电导特性。这个特性与绝缘体和金属导体之间的关键分界点有关。瓦泰和同事们写道:“这一发现表明,生命物质中普遍存在着一种电荷传导机制。”虽然他们的发现无法证明量子怪异性能解释生命机制,但他们的确让我们意识到,由量子调节的大分子领域确实有可能是我们见证全新物理学诞生的地方,而那种物理学正是薛定谔以及他那一代人所期盼的。
思想的碰撞
理论物理学家约翰 • 惠勒(John Archibald Wheeler)常说,重大科学进展更多时候起源于思想的碰撞,而非事实的稳定堆积。生物物理学正处于两大科学领域的交汇之处:物理科学和生命科学。这两个领域不但有各自的语言,而且有各自鲜明的概念框架。物理学的根基是力学概念,而生物学的根基则是信息概念。由此产生的思想碰撞预示着生物物理学必然是一片全新的科学前沿阵地。在这个领域内,现在已经被正式视为物理量(或者说一组物理量)的信息占据了核心位置,且起到了整合物理学与生物学的作用。
过去几十年内,分子生物学的巨大进展很大程度上应该归功于力学概念在生物系统中的应用,也就是要归功于物理学在生物学领域的渗透。神奇的是,现在发生的过程正好与此相反。许多物理学家,尤其是那些在量子力学基础问题领域耕耘的物理学家,都提倡把信息放到物理学的核心位置。另有一部分物理学家则猜测,新物理学潜藏在惹人注目却又令人困惑的生物有机体世界中。生物学正逐渐成为物理学的下一个伟大前沿阵地。
资料来源Physics Today
____________________
本文作者保罗 • 戴维斯(Paul Davies)是亚利桑那州立大学坦佩分校物理系教授,也是该校科学基础概念中心主任。